西安交大科研人员在神经元胞吐—胞吞的偶联平衡机制方面取得重大进展
突触传递是神经元之间进行信息交流的基本方式,神经元的兴奋信息以不同的动作电位发放模式进行编码,动作电位到达突触前膜引起突触前膜去极化,Ca2+内流,进而触发囊泡融合(胞吐)与神经递质分泌。胞吐之后,网格蛋白介导的胞吞(clathrin-mediated endocytosis, CME)、巨胞吞(bulk endocytosis)等多种胞吞模式协同作用与胞吐过程紧密偶联,以实现囊泡膜成分与膜蛋白的快速回收和再利用,这对维持细胞膜的动态平衡与神经递质的可持续分泌至关重要。作为触发囊泡与质膜融合的信号分子,大量研究表明Ca2+还是触发和加速胞吞的关键分子,仍有许多研究表明Ca2+还可抑制胞吞但其机制不明,这使得Ca2+对胞吞的调控作用备受争议,成为本领域40年来的重要谜团。
突触结合蛋白Synaptotagmin 1 (Syt1)作为起始囊泡融合的Ca2+感受蛋白与分子开关,与Complexin一起协同控制着SNARE蛋白复合体的构象变化,进而起始囊泡与质膜的融合。除响应Ca2+直接触发囊泡分泌外,Syt1还参与了囊泡的锚定与激活过程,并可能通过与Stonin 2、AP-2等内吞蛋白的相互作用参与胞吞的调控过程。然而,Syt1如何响应Ca2+参与胞吐-胞吞的偶联平衡调节,在不同神经兴奋模式下不同胞吞模式之间如何转换尚不清楚。
西安交大生命学院王昌河教授课题组系统研究了Syt1对胞吞过程的调控作用,他们首先采用对pH敏感的绿色荧光蛋白pHluorin标记突触囊泡蛋白Synaptophysin(Syp-pH)对神经突触的胞吐-胞吞过程进行实时成像(图1),发现Syt1对胞吞过程具有双相调控作用,可同时抑制快速相胞吞和促进慢速相胞吞,以确保胞吐-胞吞精密的偶联与平衡。
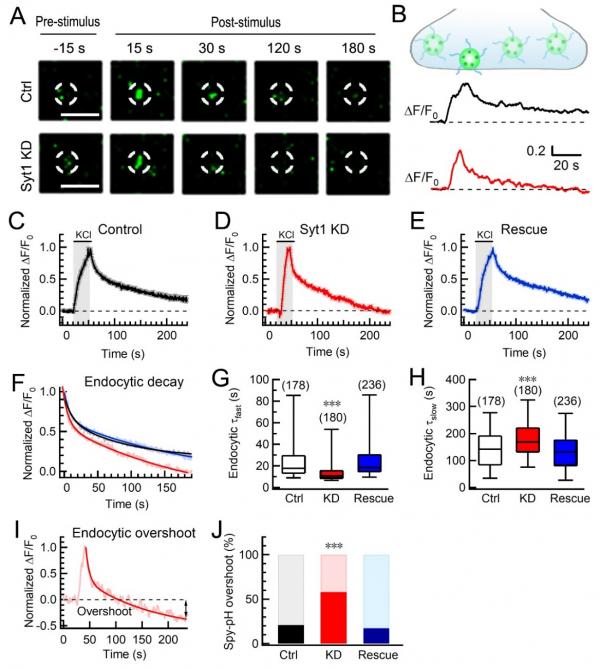
图1 突触传递过程中Syt1对胞吞的双向调控作用
为进一步研究Syt1对不同胞吞模式的调控作用,研究人员在神经元中表达Syp-pH来标记突触囊泡(含分泌囊泡与内吞囊泡),通过dextran uptake标记内吞囊泡,采用超高分辨(40 nm分辨率)STED成像观察内吞囊泡的大小和数量(图2),发现Syt1可促进直径较小的慢速相胞吞过程(CME)而抑制大直径的快速相胞吞(巨胞吞)过程。同时,采用Syt1不同突变体对KD神经元的挽救实验表明,Syt1 C2A和C2B结构域均可Ca2+依赖性地促进CME过程,但Syt1对巨胞吞的抑制作用严格依赖于C2B结构域与Ca2+的结合。
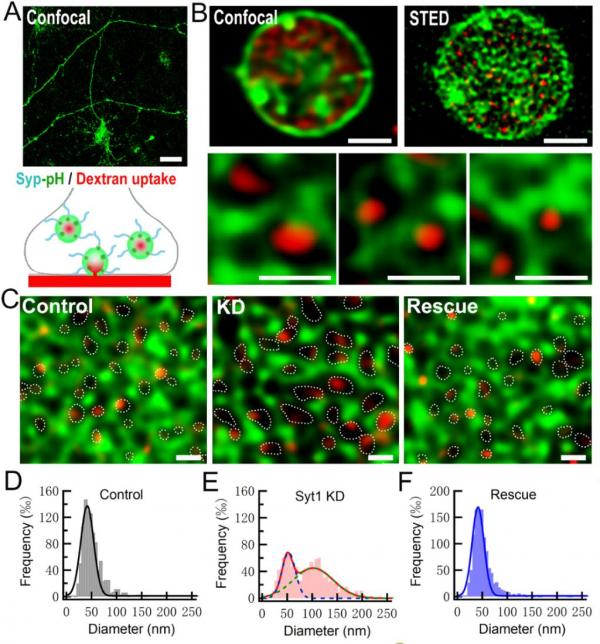
图2 Syt1敲低促进大内吞囊泡的形成
研究人员通过体外脂质体形变实验还发现,Syt1可Ca2+依赖性地促进脂质体生成脂质小管,但存在粗、细两种不同的脂质体小管(图3),而在无Ca2+条件下、或去除C2B结合Ca2+能力的突变体则只能促使脂质体产生粗管,而不能产生细管,表明Syt蛋白可介导脂质体发生两阶段的“弯曲形变”,而C2B结构域与Ca2+的结合是Syt1促进粗管向细管转变的关键所在,可能是促进巨胞吞模式向CME转变的分子基础。
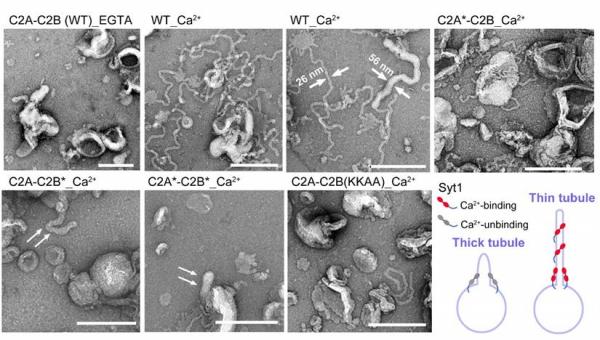
图3 Syt1介导脂质体的“两阶段”弯曲形变
综上,该研究证明Syt1可作为Ca2+感受器在神经突触中促进CME以确保高效的膜回收,同时抑制巨胞吞以防止过度胞吞(图4),不仅发现Syt1对巨胞吞过程的抑制作用,还首次揭示了Ca2+对胞吞的抑制机制,终结了本领域长达40年之久的Ca2+对胞吐-胞吞偶联平衡双向调控的争议,并提出两阶段膜重塑模型,为不同胞吞模式之间的转换机制提供依据,并为其它膜融合与膜分裂事件的调控机制提供参考。
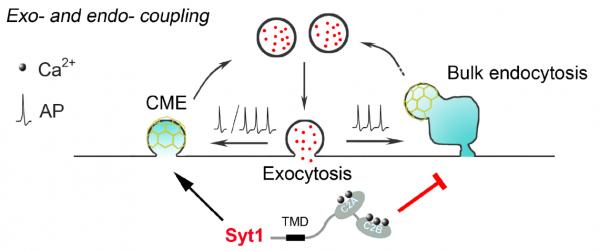
图4 Syt1促进CME抑制巨胞吞作用模式图
该研究成果以《突触结合蛋白Syt1作为Ca2+感受器双向调控神经元胞吞》(Synaptotagmin-1 is a bidirectional Ca2+sensor for neuronal endocytosis)为题在线发表在《美国科学院院报》(PNAS)上。西安交通大学生命学院王昌河教授、北京大学柴祖映博士和黄荣博士为本文共同通讯作者。博士生陈洋、胡绍琴、吴轩昂和谢振丽为本文共同第一作者。西南医科大学康新江教授和徐华栋副教授等参与了该项研究,西安交通大学分析测试中心郝英工程师协助完成STED成像数据的采集,北京大学生命学院公共仪器平台胡迎春博士协助完成电镜数据的采集。该研究得到国家自然科学基金、陕西省科技创新团队项目、国家科技创新项目、四川省自然科学基金和中国博士后科学基金的资助。
原文链接:https://www.pnas.org/doi/10.1073/pnas.2111051119
 国家重点研发计划 “质谱仪用分离打拿极电子倍增器研制”项目启动会暨实施方案论证会召开
国家重点研发计划 “质谱仪用分离打拿极电子倍增器研制”项目启动会暨实施方案论证会召开 学校召开基础学科拔尖学生培养计划2.0基地建设工作推进会议
学校召开基础学科拔尖学生培养计划2.0基地建设工作推进会议